
The Allee effect is familiar to anyone working in conservation, often colloquially described as the phenomenon whereby at low densities, populations become more vulnerable to extinction. This contradicts one of the assumptions of basic population models, which is that when competition for resources is low, populations should grow quickly. Instead this advantage is overcome by other factors, such as the difficulty in finding mates, or in resisting predation.*
More strictly, Allee effects are defined as positive density dependence in populations, that is to say, where increasing population density actually increases the fitness of individuals. When Allee effects are strong, they can result in populations shrinking in size when they’re below a critical level known as the Allee threshold. Above this point, the population will grow until eventually competition takes over and it reaches its carrying capacity (the equilibrium at which births and deaths are exactly in balance). Below the Allee threshold, the population shrinks and will inevitably go extinct. In conservation this is a bad thing, but if you’re trying to control an invasive species or a crop pest then it can be very helpful. Allee effects are therefore extremely important in applied ecology.
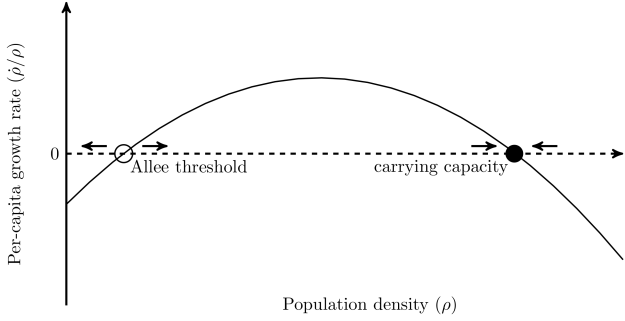
The growth rate of populations varies with their density. At high densities competition for resources causes the population to settle on a stable equilibrium known as the carrying capacity. At low densities the population slips beneath the Allee threshold and starts shrinking.
That’s how it works in theory. But in my research over the last few years, working with Jorge Velazquez, we’ve been examining how these simple population models respond when you take into account the spatial patterning of populations. Most individual organisms are not distributed regularly across habitats. Instead, they are often clustered, which means that from the perspective of an individual, the actual population density it experiences is higher than the average for the habitat as a whole. If individuals are spread out (perhaps in territories) then the opposite will be true.
This becomes important because species vary in the range over which they can do important things such as mate or disperse their offspring. A tree such as silver fir is wind-pollinated, and therefore effectively unlimited in the distance over which it can mate with other trees. Its seeds, however, are large and don’t travel anywhere near as far. In other words, it mates over longer distances than it disperses. Other trees, such as dipterocarps, are pollinated by insects which are unable to fly very far, and also have massive fruits that mostly fall right next to their parents. They are limited in both mating and dispersal.
Jorge and I had the idea that this difference in the ranges over which individuals could mate or disperse might affect their vulnerability to Allee effects. The missing element, however, was finding a species that could disperse over long distances but only mated over a short range. I couldn’t think of a tree with those characteristics**, but another organism I’ve worked on was ideal — barnacles!




In our new paper in Ecological Modelling we show that this variation in relative ranges of mating and dispersal changes the behaviour of whole populations, and makes some species more sensitive to Allee effects than others. We first show the principle mathematically, then demonstrate it using models for each of the three species above.
Fir trees don’t have any particular problems at low densities, although once populations build up they compete strongly for space because they can’t disperse their offspring very far. Dipterocarps, on the other hand, benefit from being clustered, because this makes it more likely that they will be able to find a mate.*** Their Allee threshold goes down; in other words, they are more tolerant of low population densities, and even of high mortality rates, as might occur if there is harvesting of trees. This benefit occurs despite competition for space within clusters.
Barnacles are an odd case because, although they don’t move during their adult life, their larvae are widely dispersed in the water. Nevertheless, barnacle larvae don’t just wash up on rocks randomly. They decide which areas to settle in, and once they find a suitable location, they move to be closer to other barnacles. In other words, barnacles deliberately cluster. This gives them the best of both worlds: they can escape competition from their parents, but benefit from the physical proximity required to reproduce. Their Allee threshold drops even further and their populations are highly resilient.
Measuring the ranges over which species can mate or disperse can have important implications in conservation and applied ecology. It’s not just a matter of having more accurate models; these principles could be used to identify species with particular combinations of traits which cause them to be vulnerable to Allee effects, and thereby make conservation of rare species more effective. Our models show that when finding a mate is the greatest challenge faced by an organism, increasing their clustering boosts the resilience and persistence of their populations. This trick might turn out to be very useful.
Velazquez-Castro J & Eichhorn MP (2017). Relative ranges of mating and dispersal modulate Allee thresholds in sessile species. Ecological Modelling 359, 269–275.
* It’s sometimes said that random fluctuations in small populations (e.g. in sex ratio) that increase their probabilities of extinction are Allee effects. I don’t agree that demographic stochasticity should be included because it doesn’t alter individual fitness.
** Please let me know if you can! Perhaps there’s a species with tiny pollinators but which is animal-dispersed. My expectation is that this should be a very rare combination of traits because their populations would be very unstable, and I’d be interested to see if any species can manage it.
*** If you’re wondering whether this might cause inbreeding, then that’s a reasonable question, but the answer isn’t straightforward. There is some evidence that species with poor pollen dispersal are more tolerant of inbreeding, which would reduce the apparent costs. There might be a complex evolutionary relationship between dispersal mode and inbreeding tolerance which is something to consider another time.



